
Scions of Sahul: the steadfast Australian settlers who held off sedentarism for 45,000 years
Tales from a weird continent: on Mega Marsupial slayers, the many races of Denisovans, etc.


Australia is the smallest of the seven continents, with less total area than either the US or China. And only Antarctica has less biological productivity; over 70% of the Australian landmass receives under 20 inches of rain per year, its vast core given over entirely to desert. Australia’s highest mountain registers 7,300 feet tall, barely topping the highest of the worn-down, ancient peaks in the eastern US’ Appalachians. But what the smallest continent lacks in geological dynamism, it has always lavishly made up for in biological oddities. When the first platypus specimen arrived in Europe in the late 1700’s, naturalists at the British museum trusted it was a hoax and set about searching for stitches in the bizarre chimera. Of course, the platypus is genuinely the representative of a unique egg-laying mammalian lineage, the monotremes, of whom the hedgehog-like echidna is the sole other living example.
But the mammals most associated with Australia are of course the marsupials, distinct from the placental species who reign supreme on every other continent. Unlike all other mammal offspring, marsupial young emerge at a very undeveloped stage, naked and blind, consigned to complete their maturation in a pouch. Though today their diversity peaks within Oceania, Australia’s marsupials actually originated in South America, where over 100 species of opossum continue to flourish. Both fossils and genetics confirm that the staggering variety of Australian marsupials descends from a single South American lineage, a distant relative of the Chilean colocolo opossum, which is presumed to have traversed late Mesozoic Antarctica, in T Rex’s time, before reaching Australia.
There are exceptions to marsupial supremacy in Australia. The continent is home to some 75 species of bat and 60 species of rodents closely related to mice and rats. But the biggest existential threats to marsupial mammalian hegemony are two latecomers to the continent: humans, who appeared tens of thousands of years ago, and dingoes, who arrived just 4,000 years ago. The dingo, a feral Southeast-Asian dog that likely hitched a ride via Austronesian boat to northern Australia’s shores, explains the disappearance of marsupial predators like the Tasmanian Tiger. But its impact pales next to the incursions of Homo sapiens, who fatefully transformed Australia’s ecological landscape and faunal diversity. When ancestors of the Aboriginal Australians arrived beginning 45,000 years ago, they promptly drove the vast majority of marsupial megafauna to extinction. Their interference paved the way for kangaroo species to dominate the continent’s interior today; they literally cleared the field of kangaroo competitors, their frequent woodland burnings shaping environments friendly to the now ascendant species.
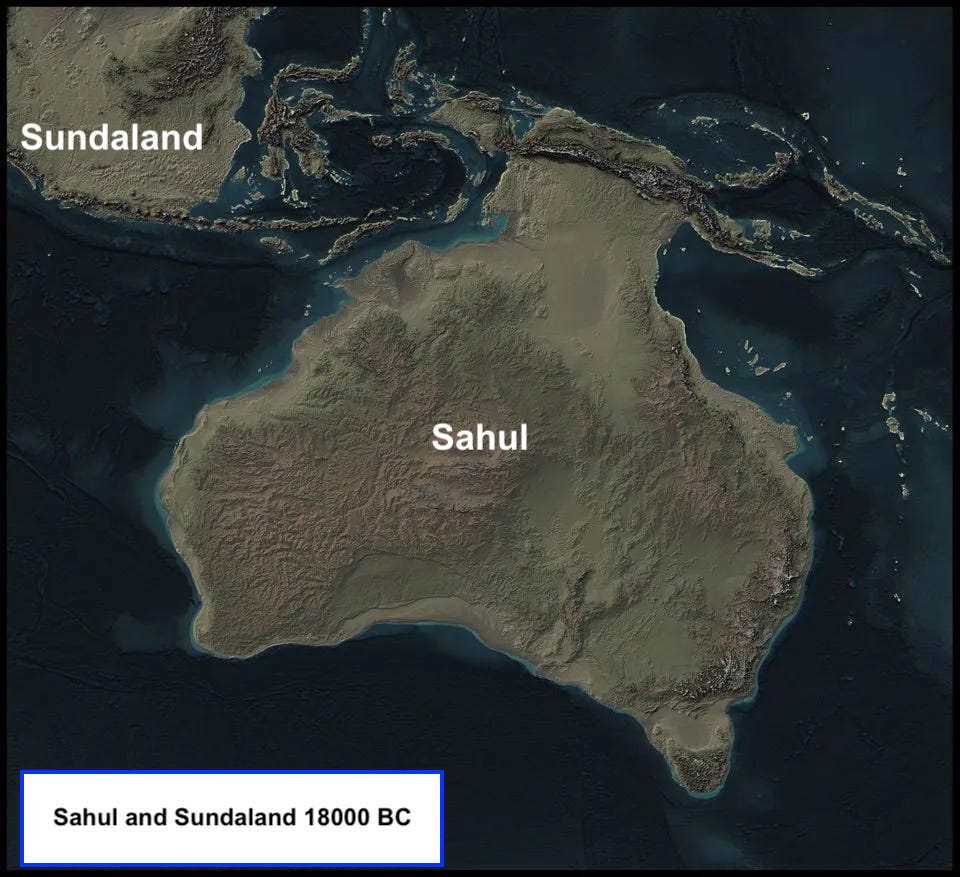
But modern human migration into Australia is but one thread in a story that touches far more than just the ecology of the planet’s smallest continent; it is a piece of the broader narrative of the hominin push past the edge of the Indo-Malaysian biogeographic zone. Here, on Eurasia’s southern fringe, familiar Asian species like tigers, elephants and orangutans, give way to New Guinea and Australia’s kangaroos and other marsupials. The genus Homo were present in Indonesia as early as a million years ago, and Flores Hobbits launched tentative explorations beyond the Wallace Line that divides the biogeographic zones with Eurasia on one side and Australia and New Guinea on the other. But those earlier human populations never seem to have penetrated deep into the Australian biogeographic zone, which during the Pleistocene included the lost continent of Sahul, plus Australia, New Guinea and Tasmania, all interlinked by land bridges that emerged as sea levels receded. The last human breakthrough to Sahul was the work of our lineage of modern humans, as they crossed the open ocean between Asia and Australia, ending the hegemony of marsupial megafauna.
In the process of Sahul’s settlement, the Australian Aboriginals’ maritime ancestors traversed one of the apes’ most speciose ecoregions: Southeast Asia, where gibbons, siamangs and orangutans coexisted not only with Homo erectus, but also with the Hobbits, Homo floresiensis, another hominin in the Philippines, Homo luzonensis, and finally, disparate populations of Denisovans. It was here at the end of the human world that over 45,000 years ago ancestors of the Australo-Melanesians, of whom the Australian Aboriginals are the most well-known, began their most impactful interactions with our cousins on Southeast Asia's frontiers. These forays would result in entanglements whose rediscovery millennia later would rock our understanding of human evolution, because as these pioneering ancient mariners embarked on humanity’s most robust experiment as nomadic foragers, they were charting a course straight for long-lost cousins who had left Africa more 600,000 years prior.
The third humans
Neanderthals were always unique in the firmament of hominin species because, unlike Homo erectus, or the recently discovered Flores Hobbits, debate long swirled over whether they were truly a distinct species from ours. We saw they were different, but had to acknowledge their eerie similarity to us, as evidenced by everything from their production of art to burial rituals; they were a sister lineage to anatomically modern humans, and some proponents of multiregional evolutionary models even argued there was substantial Neanderthal ancestry in modern Europeans. This is the major reason that geneticists’ 2010 discovery of Denisovans was so shocking: here was a sister lineage to Neanderthals that had flourished in eastern Eurasia, but which had wholly slipped under paleoanthropologists’ radar. For decades, scientists vigorously debating the relationship between two very similar species of humans, ourselves and Neanderthals, had remained oblivious to the reality that it had in fact been a crowd of three. Denisovan remains have surely languished in museums right under our noses all along, but the political and social tumult in East and Southeast Asia in the 20th century may have hampered their identification. Today some researchers believe that fossils like “Dragon Man” are actually Denisovan. Until we can get DNA from a very intact skeleton that allows us to triangulate morphological characteristics with DNA, we are left with possibilities rather than probabilities.
Aside from the unexpected fact of their very existence, perhaps the most shocking thing about the Denisovans, first discovered from a single genome in Siberia, is that the modern population with the most detectable ancestry from these prehistoric humans today are Papuans, the indigenous people of New Guinea, 5,500 km from Denisova cave, beyond China, Thailand and Indonesia, in the far-flung lands bridging the Indian and Pacific oceans. Our understanding of the geography of Denisovan admixture into modern populations has seen refinements since 2010, thanks both to two other Denisovan genomes and the massive progress generally in whole-genome sequencing of humans. In 2010, when Denisovans were discovered, on the order of thousands of human genomes had been sequenced; today there are millions. We now know that Denisovan ancestry is found across Asia, with proportions around 0.2% in South Asia, 0.1% in East Asia (and among Amerindians), and pockets of even slightly higher levels in Indonesia. The current best estimate is that 3% of modern Papuan ancestry comes from Denisovans. A similar proportion holds for indigenous Australians and other Melanesians. This is a function of their common ancestry from an ancestral population that mixed with Denisovans and subsequently diversified. Curiously, at 4%, the Negrito people of the Philippines, and in particular the Agata tribe, have the most Denisovan ancestry of any human population, pointing to a distinct, diverged demographic trajectory from the Australo-Melanesian people to their south and east.
Fascinatingly, variations in Denisovan ancestry distribution across modern humans can be observed to stand in stark contrast to the simplicity of our Neanderthal inputs. Non-African populations (representing 90% of humans alive today) range between roughly 1.5-2.5% Neanderthal ancestry, with that heritage clearly all dating to a single admixture event some 50,000 years ago. The variations today between those 1.5% and 2.5% values reflect disparate inputs of later admixture from populations without Neanderthal ancestry; those contributions then variously diluted the long-standing Neanderthal signal in western Eurasia. In contrast, the high variability in Denisovan ancestry suggests human ancestors in the east were subject to multiple admixture events, and closer inspection of the Denisovan genes in these modern human populations also makes plain that they carry traces of multiple distinct Denisovan populations.
The Denisovans sequenced thus far are Siberian, and that population’s DNA is naturally much closer to the Denisovan-admixture in East Asians and Amerindians, as opposed to the Denisovan admixture in South and Southeast Asians or Australo-Melanesians. The diversity of the Denisovan genes shows that unlike Neanderthals, Denisovans were a diverse and structured population whose distinct branches mixed with different human populations at different times. While the Neanderthals, their homogeneous population of western cousins occupied a band of frigid territory from the Atlantic to the Altai, tundra and boreal woodland, Denisovans ranged across Eurasia’s entire eastern half, from the tundra in the north to Sundaland’s tropical forests in the far south. The continent of Sundaland sank with rising sea levels, its highest points remaining as the fragmented archipelago we know as Indonesia. When it was Sundaland, tropical woodlands predominated, offering perfect human habitat. Assembling the genetic variation in the fragments present in modern humans, we can see that the genetic divergence among Denisovan populations was often as great as, for example, that between Khoisan foragers and the rest of humanity today, which represents a split on the order of 200,000 years ago. So we can think of Neanderthals, with their small populations and homogeneity, as one “race,” while Denisovans were plainly many.
By comparing the genetic distance between the individual prehistoric sequenced Siberian Denisovans and modern humans’ various quotas of Denisovan DNA segments, researchers can assess how many different Denisovan populations mixed into a given target group. East Asians today, for example, exhibit two discernibly distinct Denisovan inputs, one nearly identical to the Siberian Denisovans sequenced, and another that more resembles the Denisovan detected in Southeast Asia populations. A plausible hypothesis for the origin of this specific pattern is that as modern humans pushed into Southeast Asia, they mixed with one Denisovan population. These modern populations had a bit of Denisovan ancestry and migrated northward where they encountered a second Denisovan population, the Siberian branch, eventually mixing with them as well.
Drilling down deeper into Australo-Melanesian DNA also yields at least two Denisovan admixtures in their genomes, neither of which matches the Siberian Denisovans. Considering the likely path of migration eastward out of Africa, these admixtures would be from Southeast Asian Denisovan lineages. The genetic data also suggest that those brushes with each distinct Denisovan group fell some 15,000 years apart. The population of the older signature contributed Denisovan ancestry to both mainland Southeast Asians and Australo-Melanesians. Meanwhile, the second population only contributed ancestry to Papuans and their relatives. So the elevated Denisovan admixture in Australo-Melanesians is a compound of two elements acquired at different places and times. Denisovan segment length and distribution in the genome suggests the western Denisovan admixture occurred about 45,000 years ago, shortly after admixture with Neanderthals in the Middle East (this latter is shared universally across non-African lineages today), while the eastern admixture occurred some 30,000 years ago. By then, when modern humans had come to dominate the entire Old World, long since our lineage’s explosion was first sparked in Northeast Africa around 50,000 years ago, these last Denisovans would have been relics of a bygone age of hominin diversity.
The latter estimate implies that Denisovans may have outlasted Neanderthals by a good 10,000 years, sharing the planet with our own lineage almost until the Last Glacial Maximum 27,000 years ago. Furthermore, the easternmost Denisovans’ more restricted admixture input only to Australo-Melanesians implies that these latter may actually have migrated beyond the Wallace Line, perhaps even right up to Sahul’s edge. We know modern humans were not the first population to ford the open ocean and cross the Wallace Line. The diminutive Flores Hobbits’ ancestors crossed 20 kilometers of uninterrupted ocean, so such a feat seems plausibly within the Denisovans’ capabilities.
The precise details of how the people of Melanesia and Australia acquired their Denisovan admixture is still being reconstructed, but the attested existence of this ancestry goes a long way towards explaining peculiarities in the genetic estimates for how these peoples relate to other non-Africans. Existing data should have long suggested Denisovan admixture to us, but the paradigms had no room for it, so it hid in plain sight, assumed to be a statistical anomaly, summarily ignored. Results that implied Oceanians were more genetically distant than they should be in a simple model of demographic diffusion out of Africa, or gene variants that seemed too distinct and diverged, were shrugged off.
But archaic hominin ancestry is not even the Australo-Melanesians’ only paleoanthropological mystery. We have yet to understand how and when modern humans themselves reached Sahul, and finally settled Australia and New Guinea.
