
Science giveth and science taketh away
Reading the evolutionary book of life in 2025


The more one thinks, the more one feels the hopeless immensity of man’s ignorance.
–Charles Darwin
Not even two full centuries have passed since Charles Darwin reluctantly lobbed his meticulously built grenade into Victorian society, averring in On the Origin of the Species, that “If it could be demonstrated that any complex organ existed, which could not possibly have been formed by numerous, successive, slight modifications, my theory would absolutely break down. But I can find no such case.” In the intervening 166 years, not only has natural science gone from the almost exclusive pursuit of leisured English gentlemen like Darwin, to a complex, industrialized global knowledge production system churning out reams of data and theory by the minute, its millions of subsequent practitioners still have yet to “find such [a] case.” Darwin’s first-draft theories have proven almost miraculously robust and been profitably extended to illuminate both our deep past and our present.
Darwin’s insights marked a great leap forward for scientific theory. And the suite of fertile fields they spawned have continued to chart periodic great leaps forward ever since, including a truly miraculous one still unfolding today. Consider this discontinuity, though. Darwin could sit alone in his Down House study, mulling over the fruits of his copious correspondence with a small coterie of other curious minds, and indeed perhaps he alone was poised to not just posit evolution by means of natural selection (as Alfred Russell Wallace had done independently, spurring Darwin to set aside his book project and rush a paper out first to avoid being scooped) but to advocate for the nascent theory in volume after volume, beginning with the Origin of the Species. Today, truly no mind, however singular, works so independently.
Science since Darwin has attained the distributed complexity and efficiency, not to mention the staggering productivity of a thrumming beehive or anthill. Given what astounding insights we have achieved by striving in parallel, ceaselessly extending one another’s most promising innovations and insights, perhaps it is no surprise if the pursuit of scientific truth feels a little less “hopeless” today. With millions of minds trained on the same questions, the “immensity of our ignorance” still lingers, of course, but measurably, appreciably recedes with every passing year, like a great lake’s circumference contracting by the millimeter. Our efficiency has a perhaps predictable cost, though. Darwin didn’t just gather data, elaborate a theory and rush to publish his paper ahead of Wallace’s. Those competitive human behaviors and dynamics remain alive and well in the sciences. Darwin also waded in, his memorable, measured prose guiding the heated discussion and debate of the emerging theories that seized the public imagination for decades after.
Science, genomics in particular, is so productive and fast-moving today, it is a rare being who both sets their prolific lab’s pace of publishing and pauses to decode and actively mull over the implications of their findings with engaged lay readers. Every crucial interdependent part of the scientific enterprise is now compartmentalized. We have gained the world, as denominated in data, theory and deeper knowledge of life’s secrets. But along the way we have lost something. In Darwin’s age, a literate layperson could (and crucially, did) follow along with the discoveries and nascent theories of their age’s brightest minds. They could wrestle with the implications of those game-changing theories, mull over the puzzle of natural selection’s undiscovered mechanisms. And a due sense of awe and wonderment was accessible to all, thanks in no small part to Darwin’s deft and prolific popular writing on evolutionary theory as he continued elaborating it.
Today, human genetics still makes popular headlines, but the considerable ink spilled has a way of pooling on rather more prosaic adjacent questions, like whether long-term care insurers will get your 23andMe data and deny you a policy. But at this point in our society, basic assumptions about evolution, adaptation, heredity, genetics and DNA itself have all long since suffused the air we breathe. As a species, as a society, we make innumerable personal and policy decisions based on a common understanding of these concepts. Which, on balance, is all to the good. But along the way, as we benefit from all that immense scientific progress, we as lay people have lost two key things Darwin’s contemporaries could widely enjoy.
The first is simply perspective. We have summited a dizzyingly high peak today. The sweeping view our scientific accomplishments now afford us, literally in every direction, both millions of years back into our (and every species’) past, not to mention leagues deep into contemporary questions… is truly the stuff of science fiction. But a sense of wonderment, of exhilaration at what we can now understand seems almost wholly absent in the lettered public. Darwin’s suspicions about evolution and heredity apparently universally thrilled and terrified his educated contemporaries. Our advancements since then have extended his ideas light years beyond their embryonic form. And yet no commensurately eloquent champions (not to mention, obviously, originators) trumpet them to a fascinated public, let alone become household names for doing so. But perhaps humanity has outgrown its capacity for astonishment anyway. In an age when most citizens start their day by fumbling for their personal supercomputer, what could actually still wow us?
I don’t have any prescription for our general inurement to awe. But, far across the spectrum from wide-eyed wonderment lies the other gift Darwin’s contemporaries could also freely partake of. Evolution by means of natural selection, as Darwin first proposed it, lacked any mathematical formalism. It was an eloquent but simple idea, wholly elaborated in prose. Not even the elementary arithmetic ratios found in a Punnett Square of simple inheritance had yet intruded on its original formulation. In the intervening 166 years, evolution and genetics have been supercharged multiple times by powerful advances in mathematical theory and supercomputing.
And no surprise, the public conception of what evolution truly is and how genetics functions have tended to languish decades if not an entire century behind the actual state of the field. So let’s try to dig into what is truly understood in the studies of evolution and genetics today. Perhaps in the process of examining how infinitely reshuffling simple strings of just four little letters (the famous A, C, G and T) has begat Darwin’s “endless forms most beautiful,” I can share some small ration of my enduring awe at our field’s fruits. Just in this dawning millennium, the entire book of life has been thrown open for anyone to read. Let’s talk about the prosaic process of how we do just that.
Evolution is responsible for the fate of everything from the lowly virus to the grandest redwood. It is a concept most people would attest to understanding. But since Charles Darwin’s original ideas scandalized the Victorian Age 165 years ago, science has advanced almost unimaginably, and today the gap between how the public embraces and understands evolution and how scholars model evolutionary processes yawns ever wider. The 20th-century fusion of genetics with evolutionary biology, which allowed the field to transcend its reliance on natural history, at once unleashed awesome powers to project both forward and backward in time via the powerful levers of mathematical formalism. But it also balkanized research communities, with the mathematical savants and their deepest insights too often sealed off from the world of the molecular biologists at the bench and field researchers out at their sites.
Darwin’s On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life indisputably cemented his place in the scientific firmament. His work marked an epochal leap forward, but it would be a mistake to attribute this to the contention that organisms evolved from one form to another. Earlier thinkers, from his grandfather Erasmus Darwin back to the ancient Greek philosopher Anaximander, had endorsed the view that organisms changed over time. Darwin’s contribution was instead to proffer a mechanism by which evolution, and diversification into different species and lineages, might occur. That mechanism was adaptation via natural selection.
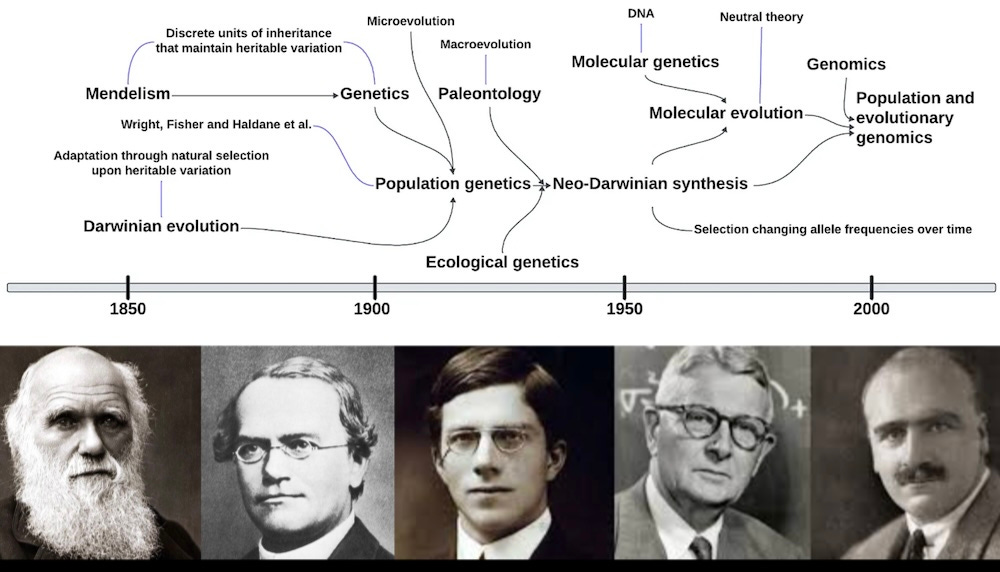
But despite its cultural and scientific impact, On the Origin of Species and its companion volumes, The Descent of Man, and Selection in Relation to Sex, were not enough to fully convince the Victorian establishment of Darwinism’s truth. A major impediment was that despite owning a volume that extensively described Gregor Mendel’s experiments (and even, apparently, having perused it enough to record some minor marginalia), whose insights would go on to form the foundation of modern genetics, Darwin never articulated a theory of inheritance that persuasively allowed for the maintenance of heritable variation, a necessary factor for natural selection to operate as a pervasive and persistent mechanism across the generations. Because, if two organisms mix their phenotypes together, naive intuition might suggest that trait by trait, successive generations would converge ever closer to the population-wide averages, erasing the very variation that natural selection requires to drive adaptive change via mixing.
The circa 1900 rediscovery of Mendel’s insight that inheritance occurred via the transmission of discrete units of hereditary factors (genes) solved the conundrum and triggered an early 20th century Darwinian renaissance. Genetics gave us a plausible mechanism of variation’s persistence across generations, genes were simply reshuffled and re-dealt across infinite successive rounds of play, without erasing the underlying unique information. With a model to test, biologists could now generate predictions. Experimentalists could now quantify the inferred variation in the laboratory. At a stroke, evolutionary scholarship bolted from the armchair and the observational garden. Darwin’s incisive intuition and copious observation were now harnessed to an engine of mathematical rigor and empirical testability. It was like taking the cosmic leap from Galileo’s telescope, as he first brought the vaguest outlines of Saturn’s rings into focus, directly to interplanetary Voyager spacecraft, beaming back vivid flyby images of the fires of Io and the dense mists of Titan.
Popular conception tends to wholly downplay the revolutionary scale of this 20th-century transformation. It is easy and common to caricature evolution as slouching apes’ rise to upright carriage and sapience, the extinction of the dinosaurs or a first fish clambering awkwardly onto land. Darwin’s theory is still cartoonishly drawn as catastrophes and barely plausible ecological miracles. But to develop an intuition for how evolution worked in the past and might work in the future, we need to conceptualize the process from the ground up; we need a “gene’s eye view.”
In the 1920’s and 1930’s, an ad hoc group of luminaries coalesced to advance and elaborate Darwin’s prescient intuitions with another great leap forward. Chief among them were R. A. Fisher, an English mathematician also instrumental in creating modern statistics, J.B.S. Haldane, an Oxford-trained mathematician and Marxist aristocrat whom James Watson called “England's most clever and eccentric biologist,” and Sewall Wright, an American experimentalist whose deepest theoretical insights were informed by his experience in the laboratory. Together, they created a mathematical framework to capture Darwinian evolution’s mechanics as a change in group-wide allele frequencies; this was the fertile germ of the field we today call population genetics. Alleles are variants of a gene. In the modern language of DNA, this usually means A’s, C’s, G’s and T’s switching around across the generations, providing the underlying biochemical substrate that encodes heritable change, complex dynamics which in the genomic age can finally be precisely tracked.
It is through DNA that evolutionary biologists can construct phylogenetic trees reaching back a billion years, detect the footprints of natural selection in the genome, and even confidently calculate that when modern humans trooped out of Africa some 60,000 years ago, the whole exodus could only have numbered some 1,000-10,000 breeding individuals. Ultimately, these analytic acrobatics draw upon the same foundation as Darwin’s theory of natural selection: heritable variation. But we don’t conjure this fuel for our genetic analyses out of thin air; heritable variation accumulates or depletes like any resource. A deeper accounting of this resource’s stores and its dynamics is critical to understanding evolution’s foundation.
The Four Forces of Variation
First of all, unlike say the speed of light in physics, genetic diversity in evolutionary models is not a fixed value or a constant. It is a protean value buffeted by evolutionary forces that remove or add variation from one generation to the next. These real forces are represented as abstract mathematical parameters, but accounting for them grants us an evolutionary time machine that can wind back into the past and project into the future. While evolution from a paleontological perspective traces the biological outputs: morphological change over time, population genetic frameworks try to grasp the process via its foundational inputs, the underlying variation of genes. These four forces are mutation, migration, selection and drift.
Most fundamental is mutation, the ultimate source of all DNA variation. In everyday parlance, mutation refers to external characteristics visible on an organism, the phenotypic deviations by which you identify “mutants” in the X-Men films. In fact, classical genetics discovered mutations in exactly that superficial way, whether white-eyed Drosophila melanogaster (fruit flies), wrinkled peas or albino mice. But today we can think of mutation in its essence as changes in DNA sequence, regardless of whether they’re phenotypically discernible. For example, point mutations like transitions between purines A↔G and pyrimidines C↔T, or transversions between purines and pyrimidines, A↔C, A↔T, G↔C, G↔T. These mutations result in single nucleotide polymorphisms (SNPs), changes away from the default “wild type” at a specific base or nucleotide that deliver variability into the population at that position; e.g., 90% A, 10% C, as opposed to a monomorphic site that population-wide turns up 100% C. Less commonly, but still significantly, mutations can be deletions and insertions within the genome, variation in copy number of a gene and even genomic rearrangements, all of which fall under the rubric of structural variation. Whatever the molecular biological detail, it is inescapable that mutation starts at the level of DNA.
Because technology now allows us to sequence DNA letter for letter across large numbers of individuals in a given species, we can also understand mutation as more than a conceptual physical phenomenon or a general mathematical abstraction; we can count changes in the DNA of any organism. Today we know the mutation rate in humans is approximately 1 × 10⁻⁸ mutations per base pair per generation. That works out to each of us living with 30 to 60 new mutations differentiating us from our parents.
If the accumulated stockpile of variation is evolution’s fuel, then mutations are like periodic explosions that obliterate mountaintops to expose rich seams of anthracite coal, thus upping the ecosystem’s aggregate available fuel. But all mutation is not created equal. Only about 1% of the human genome codes for proteins, some 30 million bases out of our three billion. And even of these 30 million protein-coding bases, many do not result in any amino acid change when mutated. In complex organisms, most mutations are mute; they don’t affect adaptation or cause biophysical change. They’re neutral to selection and nonfunctional as far as biological relevance goes. Any mutation adds heritable variation, but most of this lurks hidden away at the genomic level; selection driven by the environment or competition can only grapple with those that affect traits or behaviors.
The average human differs on about 4.5 million single SNPs from any other human, with most having no functional relevance. But in the rare cases where variation has phenotypic consequences, DNA makes the leap to classical Darwinism, leaving an unmistakable mark in the genome. For example, on human chromosome 15, at genome position 48,124,702, roughly midway along that chromosome’s some 100 million bases lies a SNP that varies across world populations, appearing as genotypes AA, AG and GG. Almost all Europeans are AA, almost all Africans are GG, and populations like Middle Easterners and South Asians fall between, preserving a local sampler of all three genotypes. Scientists discovered this mutation, at the gene SLC24A5, in a small fish species, where it causes depigmentation. In humans too, it strongly correlates with skin color. This is variation that matters to selection, the sort of variation that can drive adaptation.